Õisikute ilmumise bioloogiline tähendus on nii entomofiilsete kui ka anemofiilsete taimede õite tolmlemise tõenäosuse suurenemises. Pole kahtlust, et putukas külastab ajaühikus palju rohkem õisi, kui neid koguda õisikutesse. Lisaks on roheliste lehtede seas rohkem märgata õisikutesse kogutud lilli, mitte üksikuid õisi. Paljud rippuvad õisikud õõtsuvad õhu liikumise mõjul kergesti, hõlbustades seeläbi õietolmu levikut.
Õisikud on iseloomulikud valdavale enamusele taimelilledest. Tavaliselt on õisikud rühmitatud taime ladva lähedale okste otstesse, kuid mõnikord, eriti troopilistel puudel, ilmuvad need tüvedele ja jämedatele okstele. Seda nähtust tuntakse kui caulifloria. Näiteks on šokolaadipuu. Arvatakse, et tingimustel troopiline mets Lilllill muudab lilled tolmeldavatele putukatele ligipääsetavamaks. Teine näide lilleõiest on kaunviljas Cercis carob, mida kasvatatakse laialdaselt Krimmis ja Kaukaasias.
Õisikul on põhitelg ehk õisikutelg ja külgteljed, mis võivad olla erineval määral harunenud või hargnemata. Nende otsaoksad - pedicels - kannavad lilli.
U lihtneÕisikute külgteljed ei ole hargnenud ja on varred. Nimetatakse õisikuid, milles hargnevad külgmised teljed keeruline. Komplekssel õisikul võivad olla esimese, teise ja järgneva järgu külgmised teljed.
Peamised ja kõik otsmised külgteljed võivad lõppeda apikaalsete õitega, mille tagajärjel on nende kasv piiratud. Selliseid õisikuid nimetatakse suletud ( sümpodiaalne) või teatud. Suletud õisikutes avanevad tipuõied tavaliselt varem kui all olevad külgmised ja seetõttu nimetatakse neid tipuõiteks.
Avatud õisikutel on peatelje kasv piiramatu (s.t. lahtine) ja õied paiknevad õitsva võrse morfoloogilise tipu küljel. Sama võib kehtida ka külgtelgede kohta. Selliseid õisikuid nimetatakse avatud ( monopodiaalne) või määramata. Avatud õisikutes õitsevad õied järjestikku alt ülespoole, mistõttu nimetatakse neid külgõitsejateks.
2. Monopodial või botryodnye õisikud.
Ühejalgsetel (avatud või külgõieliste) õisikutel on peatelg, mis jätkab kogu aeg kasvu, ja külgteljed kannavad õisi. Sellises õisikus puhkevad kõigepealt alumised õied ja seejärel järjest ülemised. On lihtsaid ja keerulisi botrüoidseid õisikuid
Lihtsad botrüoidsed õisikud
Pintsel - peateljel või esimest järku teljel asetsevad vaheldumisi pedicellid, mis kannavad väikseid lilli (maikelluke, linnukirss, rebashein);
Kass on õisiku peateljeks, erinevalt ratsapuust ripub ta allapoole (kask);
Spike – piklikul peateljel on istuvad lilled (jahubanaan);
Kõrv - õisiku põhitelg eristub suure lihakusega, õite asetus on kõrva (maisi) oma;
Sh i t o k - õied asetsevad samas tasapinnas, alumised õied on pikemate varrega kui ülemistel (pirn, õun, ploom);
Vihmavari - üksikute lillede varred on ühepikkused ja ulatuvad telje ülaosast nagu vihmavarju (sibul, kirss) kodarad;
Korv - arvukad istuvad lilled asuvad lühendatud ja taldrikukujulisel paksenenud teljel. Väljaspool on kandelehtede ümbris (päevalill, kummel);
Pea – tihedalt asetsevad lilled (ristik, burnet) istuvad lühendatud ja laiendatud peateljel.
Komplekssed botrüoidsed õisikud
Neid iseloomustab asjaolu, et lilled paiknevad 2., 3. jm järgu telgedel.
1. Panicle – külgtelgedel on lihtsad harjad (sirel, viinamarjad).
2. Kompleksne oga – peateljel paiknevad lihtsad ogad (nisu, rukis, oder).
3. Kompleksvihmavari – külgtelgedel on lihtsad vihmavarjud. Lihtsatel vihmavarjudel on tavaliselt oma kandelehed (privaatsed involucres) ja vihmavarju põhjas olevad kandelehed moodustavad ühise involucre (till, porgand, aniis).
4. Kompleksne korümb (corymbose panicle) - külgtelgedel on väikesed õisikud-korümbed (pihlakas).
Mõnel juhul täheldatakse segatud õisikuid. Näiteks kaeral on õisik õisik, kuid õite asemel on väikesed ogakesed, nii et kaks õisikut on segunenud: õisik ja oga.
3. Sümpodiaalsed või tsümoidsed õisikud. Puuviljade morfoloogia.
Sümpodiaalsed (cymoid) on õisikud, mille harilik vars (peatelg) lõpeb õiega ja selle kasvu jätkavad ülemised külgvõrsed ehk külgvõrsed, mis omakorda jätkavad oma kasvu samal viisil. Tsimoidsed õisikud hõlmavad järgmist:
1. Hark ehk dihhaasia. Peatelg lõpeb õiega, varre kasv jätkub all olevate vastaskülgsete pungadega, moodustades õisi jne (nelk, kukeseen, seebileht).
2. Pleiochasium ehk valevihmavari – varre kasv lõpeb õiega ja jätkub õisi moodustavate külgmiste pungade all keerdudes (euphorbia).
3. Curl - varre kasv lõpeb õiega ja jätkub külgpungaga, mis moodustab õie ja nii korduvalt, kõik õied on suunatud ühte suunda (kartul, unustamata, harilik hari).
Puuviljade morfoloogia.
Pärast viljastamist arenevad pesa munasarja seinad viljakesteks (perikarp). Perekarp on seemet ümbritsev viljasein, mis moodustub samaaegselt viljakestaga. Perikarp koosneb tavaliselt kolmest kihist:
1) väline ( eksokarp), mõnikord kaetud erinevate väljakasvudega (vahtra lõvikala, takjahaagised);
2) keskmine ( mesokarp), mis moodustab mahlaste puuviljade viljaliha, mis sisaldab palju suhkrut (ploom, kirss) või õli (oliiv);
3) sisemine ( endokarp), sageli muutudes mahlaste puuviljade kihiks
kivisteks rakkudeks (ploomi, kirsi süvend) või mahlakaks viljalihaks (sidrun).
Kõik kolm tsooni on selgelt eristatavad. Näiteks kirsi viljal on õhuke nahkjas väliskiht - eksokarp, vilja söödav mahlane viljaliha on mesokarp, üksikut seemet ümbritsev kivisest koest kõva kivi - endokarp.
Valmimata viljad on tavaliselt rohelist värvi, eksokarp- ja mesokarprakud sisaldavad klorofülli ja fotosünteesivad. Vili küpsedes kaotab oma rohelise värvuse ja omandab sageli erksa värvuse, mille põhjuseks on kas rakumahlas leiduv antotsüaniin (kirss, viinamarjad, mustikad) või kromoplastid (pihlakad, tomat).
Mõnel taimeliigil moodustuvad viljad mitte ainult pesa munasarjast, vaid ka anumast või periantist. Selliseid vilju nimetatakse valedeks. Näiteks maasika marjakujuline valevili on ülekasvanud mahlane värviline anum, millel on väikesed kuivad päris viljad (achenid), mis on moodustunud pesade munasarjadest. Ülekasvanud anumast moodustuvad kibuvitsa-, õuna- ja pirnide viljad.
Mõnel taimeliigil moodustub vili mitmest pesast. Sellist vilja nimetatakse kokkupandavaks või kompleksseks (vaarikate, murakate, tibude liitvili).
4. Puuviljade klassifikatsioon. Kõik puuviljad on jagatud järgmisteks tüüpideks.
Kuivatatud puuviljad
1. Pähklil või pähklil on puitunud viljakest, mis sisaldab ühte lahtist seemet. Näiteks sarapuu, tamme, pöögi, kanepi puhul.
2. Ahenel on nahkjas viljakest, mis sisaldab ühte vabalt lebavat seemet. Näiteks päevalillel, võilillel, kummelil ja teistel Asteraceae perekonna liikidel.
3. Karüopsisel on nahkjas viljakest, mis sisaldab ühte seemet, mis on sulandunud viljakestaga. Näiteks nisus, odras ja teistes teravilja perekonna liikides.
4. Lõvikala on pähkli- või ahenekujulise viljakestaga, mis on kasvanud üheks või mitmeks tiivuliseks lisandiks. Näiteks vaher, kask, jalakas, saar.
5. Mõnel liigil on kuivanud, mittehaardunud viljal kärbes – karvatuttt. Näiteks võilill.
Kuivatatud puuviljade avamine
1. Leaflet - karbikujuline ühelokulaarne mitmeseemneline vili, mis on moodustatud ühest karbist, mis avaneb piki ventraalset õmblust tipust põhjani.
Näiteks vatiroos, ranunculaceae ja Rosaceae sugukonna liigid. Pojeng toodab ühe õie pesadest palju voldikuid.
2. Bob - karbikujuline ühekohaline, sageli mitme seemnega vili, mis moodustub ühest karpelest, mis avaneb piki kõhu- ja seljaõmblust tipust põhjani. Näiteks hernestes ja teistes kaunviljaliste sugukonna liikides. Mõnel liigil keerduvad oaventiilid avanedes spiraalselt ja puistavad seemned laiali (näiteks kollasel akaatsial). Mõnel liigil on oad üheseemnelised (näiteks esparseen) ja siis ei avane.
3. Kaun ja silique on karbikujuline, kahelokaalne, mitme seemnega vili, mis on moodustunud kahest karbist. Vilja keskel ülalt alla on kilejas vahesein, mille äärte külge kinnituvad seemned. Vili avaneb kahe klapiga mööda kahte õmblust põhjast tipuni. Klapid kukuvad maha, kuid vahesein seemnetega jääb alles. Kaun on pikk ja kitsas vili – pikkus on laiusest 4 või enam korda suurem (näiteks sinep), kaun on lühike ja lai (näiteks karjase rahakott).
4. Kapsel - mitmelokaarne (harvem unilokulaarne) mitmeseemneline vili, mis on moodustunud mitmest karbist. Pungad avanevad erinevalt, kas klappidega (näiteks puuvillas, daturas, linakiust, tees, liiliates, riitsinusubades) või kukli tipus olevate hammastega (näiteks paljude nelgi perekonna liikidega), või aukudega (näiteks teatud tüüpi mooni ja kellukesega), kas kaanega karbi ülaosas (näiteks henbane'is) või külgmiste piludega (näiteks okras).
Mõnel liigil lagunevad kuivad mitmeseemnelised viljad (kaunad, oad) valmides põiki üheseemnelisteks segmentideks (näiteks metsrõigas). Selliseid puuvilju nimetatakse liigendatud.
Teistel liikidel lagunevad kuivad mitmeseemnelised viljad valmides pikisuunas eraldi üheseemnelisteks segmentideks, näiteks vahtral ja teistel vahtraliste sugukonna liikidel, samuti perekondades Apiaceae, Lamiaceae, Malvaceae jt. Selliseid puuvilju nimetatakse murdosadeks.
Mahlased puuviljad
1. Marja on mahlane, tavaliselt värviline, mitmeseemneline vili, mis on moodustunud ühest või mitmest karjast. Marjas olevad seemned on põimitud mahlasesse viljalihasse. Näiteks viinamarjad, sõstrad, karusmarjad, belladonna, mustikad, jõhvikad, kartulid, tomat, kurk, arbuus, melon, kõrvits, sidrun, mandariin, apelsin (kurk, kõrvits, arbuus, melon on valemarjad, kuna marja välimine osa lihavad viljad, mille nad anumast moodustasid).
2. Drupe - tavaliselt mahlane ja värviline, ühe või mitme seemnega vili, mis on moodustatud ühest või mitmest seemnest. Luuvilja endokarp on kivi välimusega. Näiteks ploomid, kirsid, aprikoosid, koerapuu, oliivid. Mitme kiviga luuviljad - leedris, lahtistav astelpaju. Kreeka pähkli vili on luuvili, kuid selle mesokarp ei ole mahlane; nn pähkel ise on vilja seeme. Ka mandli luuvilja mesokarp pole mahlane, samal ajal kui kookospalmi luuvili on kiuline.
Katteseemnetaimede viljakus on väga kõrge. Seega toodab üks valge kinoa taim aastas umbes 100 000 seemet, kanaliha - ≈ 500 000, kinoa - ≈ 750 000,
paplid ≈ 27 000 000 seemet.
5. Viljatus. Puuviljade ja seemnete levitamine.
Mõnel liigil moodustub vili õisikust. Seda moodustumist nimetatakse viljatuseks. Näiteks peedi vili on pall, mooruspuu vili “marja”, veinimarja vili “marja”. Mooruspuu infruktsioon on sulatatud valeviljad, mis on moodustunud ülekasvanud periantidest.
Mõnes sordis kultuurtaimed viljad arenevad ilma tolmeldamise ja viljastamiseta ning seega ilma seemneteta. Seda ilma seemneteta viljade moodustumist nimetatakse partenokarpiaks. Partenokarpiat täheldatakse mõnel õuna-, pirni- ("seemneteta"), viinamarja- (rosinad – kuivatatud seemneteta viinamarjad), karusmarjadel, mandariinidel, apelsinidel, sidrunitel, viigimarjadel, jaapani hurmadel jne. Mõne sordi puhul võivad partenokarpiat põhjustada häbimärgi ärritamine võõra õietolmuga (pirni tolmeldamine õuna õietolmuga, tomati õietolmuga, baklažaani õietolmuga), mõned kemikaalid, putukahammustused, kuuma traadiga põletamine jne. Seemneteta viljad on tavaliselt väiksemad kui seemnetega viljad ja seetõttu annavad nad väiksemat saaki. Praktikas hindavad tarbijad aga seemneteta puuvilju kõrgelt. Seemneteta sordid saavad paljuneda ainult vegetatiivselt.
Puuviljade ja seemnete levitamine
Katteseemnetaimede seemnetel ja viljadel on levimiseks palju erinevaid kohandusi. Enamik taimeliike on kohanenud tuulega seemnete ja viljade levikuga (anemokooria). Puuvilla-, haava-, papli-, paju-, võililleviljade jt seemnetel on omapärased pikad karvad ja helbed, mis hõlbustavad nende levikut tuulega, sageli pikkade vahemaade taha. Vahtra, kase, jalaka, tuha, lepa, männi, kuuse jm seemnetel on tiivulised väljakasvud (sellepärast nimetatakse vilju lõvikalaks), mis soodustavad nende levikut tuulega. Tuule levimisele aitab kaasa ka pärna õisiku katteleht. Paljude kanarbiku, emajuurte, orhideede jt sugukondadest pärit liikide seemned on nii väikesed ja kerged, et neid kannab tuul nagu tolm väga pikkade vahemaade tagant. Sfäärilise kujuga kaameli-, kurai-, putukarohu jne taimed murrab pärast viljade valmimist tuul maa-alusest osast ära ja veereb mööda steppi pikki vahemaid, moodustades sageli terveid liikuvaid šahtisid, hajudes laiali. nende seemned teel. Need taimed on üldtuntud kui tumbleweed.
Veetaimede seemned ja viljad levivad vee toimel (hüdrokoor). Vihmavesi (eriti pärast vihmasaju), ojade ja jõgede vesi mängib olulist rolli maal kasvanud taimede seemnete ja viljade edasikandmisel.
Loomadel on seemnete levitamisel suur roll. Mahlaseid vilju söövad linnud (ornitokoor) ja vähemal määral ka teised loomad (zoochory). Nende seemned, mis läbivad looma seedetrakti, ei ole kahjustatud ega kaota oma elujõulisust. Koos loomade väljaheidetega langevad seemned maapinnale ja idanevad.
Seega idanevad seemned sageli emataimest märkimisväärsel kaugusel. Mõnel taimetüübil (takjas, porgand, takjas, takjapaber) on seemned ja viljad erineva kinnitus- või kleepuva pinnaga ning kinnituvad mööduva looma karva külge või kleepuvad lindude sulgede külge ja transporditakse seeläbi. Väikesed seemned levivad sipelgate poolt ning võivad ka loomade sõrgade külge poris kinni jääda ja niimoodi kanda.
Mõned taimeliigid on välja arendanud võime oma seemneid iseseisvalt hajutada (autokoor). Nii avanevad kollases akaatsias valminud ubade klapid ja keerlevad sellise jõu ja kiirusega, et oas olevad seemned hajuvad jõuga igas suunas. Vilja selline avanemine kollases akaatsias on seletatav asjaoluga, et oa valmimisel tõmbuvad selle klappide välimine ja sisemine kiht ebaühtlaselt kokku ning nende vahel tekib ja kasvab pinge, mis põhjustab oa avanemisel klappide keerdumist ja värisemist.
“Hullas” kurgis, mille küpsed viljad murtakse varre küljest lahti ja tekkinud auku visatakse seemned koos vedela sisuga. See juhtub sellise jõuga, et seemned kukuvad tavaliselt mitme meetri kaugusele. Vedeliku ja seemnete väljapaiskumine on seletatav asjaoluga, et viljade valmimisel rõhk suureneb ja viljade küpsemise ajaks saavutab see kõrge pinge.
TUNNI TEEMA 27 : Ranunculaceae (Ranunculaceae). Mooni perekond (Papaveracea)
1. Ranunculaceae perekond. 2. Moonipere. KOOS. 379 – 383
Telli Ranunculaceae.Ranunculaceae perekond (2.000)
Umbes 2000 liiki. Ranunculaceae perekonna liigid on levinud külmas parasvöötmes ja subtroopilistes (mägistes) piirkondades. Smolenski piirkonnas kasvab 32 liiki. Troopikas leidub selle perekonna taimi erandina. Eluvormid on peamiselt mitmeaastased rohttaimed, haruldased on kääbuspõõsad ja viinapuud.
Lehed on lihtsad, ilma tähisteta; Lehtplaadi kuju on mitmekesine. Lehed võivad olla terved (kevadhein) või erineval määral tükeldatud (metsanemone, üllas maksarohi, kolumbiin). Lehtede asetus on korrapärane.
Paljudel liblikatel on maa-alused võrsed modifitseeritud (anemoonide risoom ja allika käbid selged).
Lilled võivad olla üksikud (perekond anemone) või kogutud õisikutesse (perekond ranunculus, kolumbiin, rukkilill). Ranunculaceae on õiestruktuurilt äärmiselt mitmekesine.
Lillestruktuuri mitmekesisust saab väljendada mitme evolutsioonilise seeria jälgimisega.
1. Määratlemata liikmete arvuga õied (anemoonil võib lihtpärlil olla 5-8 lehte) → kindla, selgelt väljakujunenud liikmete arvuga õied (perekonnavõitleja).
2. Lihtne pärand (perekond anemone, perekond columbine) → kaksikpärl (perekond Ranunculus)
3. karbid alates suur hulk kuni üheni
4. günoetsium apokarpsest sünkarpoosseks
5. lilled aktinomorfsetest õitest sügomorfseteni (perekond wrestler, perekond aconite
6. Lille telg on piklik, mistõttu lilleelemendid on paigutatud spiraalselt → õie telg on peaaegu tasane, mistõttu lilleelemendid on paigutatud ringi.
7. lilled on spetsialiseerimata, entomofiilsed (sageli tolmeldavad kärbsed) → õied on spetsialiseerunud, entomofiilsed (perekond wrestler, perekond Larkspur).
Paljudele võikulliliikidele on iseloomulik staminoodid- tugevalt redutseeritud ja modifitseeritud kroonlehed (või tolmukad), mis muutuvad nektariteks. Ainult basiiliku taimel pole nektareid.
Enamikul liikidel on palju püstakuid, need on vabad ja paiknevad piklikul teljel (perekonnad kõrvits, maksarohi, saialill). Harvem on püstolite arv piiratud: 3-1. Munasarjad on alati ühesilmsed ja kõrgemad. Tavaliselt on palju munarakke, kuid on liike, mille pesas on 1 munarakk. Puuviljad on voldikud või pähklid.
Tolmu võib olla erinev arv. Enamasti on lilli suure ja määramatu arvu tolmukatega, mõnel liigil võib olla 3 kuni 1 tolmukat.
raba saialill – * Р 5 А ¥ G ¥
Sõltuvalt puuvilja tüübist eristatakse Ranunculaceae perekonnas kahte alamperekonda:
· talitajad (genera ujumistrikoo, maadleja, lõoke), kellel on voldiku vilju
· Ranunculaceae (perekond Buttercup, Anemone, Basiilik), millel on pähkline vili
Alkaloidid on väga levinud kõigis selle perekonna taimedes. Ranunculaceae on mürgised, nad ei ole toidutaimed, kuid alkaloidide rohkuse tõttu kasutatakse taimi ravimid. Morfoloogiliselt mittespetsialiseerunud liikides (perekond basiilik, saialill ja võikull) sünteesitakse lihtsaid alkaloide, arenenud ja spetsialiseeritud perekondades aga kompleksseid alkaloide (taimed perekonnast aconite ja larkspur).
Alates erinevat tüüpi Buttercups sisaldab 20 kardioloogilises praktikas kasutatavat alkaloidi. Mürgised on igat tüüpi kontlillide alkaloidid, kõige mürgisemad neist, mis kuuluvad akoniitide perekonda.
Olulised ravimtaimed on kevadine adonis, larkspur reticularis ja booraks (või akoniit).
Kevadises adonises sisalduvad alkaloidid tekitavad kuraarelaadset efekti ja neid kasutatakse lihaste lõdvestamiseks.
Larkspur reticularis'e alkaloide kasutatakse närvikliinikus motoorse düsfunktsiooniga seotud haiguste korral
Radikuliidi korral soovitatakse ravimtaime Djungarian aconite tinktuuri, mis on osa ravimist "Akofit".
Haruldased ja kaitsealused liigid: euroopa ujuja, harilik vares, põhjamaadleja.
Telli Poppy.Moonipere (250)
Selle perekonna taimed on levinud peamiselt troopilistes ja subtroopilistes piirkondades. Harva leidub parasvöötmes. Smolenski oblasti territooriumil on 2 liiki - suur vereurmarohi ja moon.
Mooni - mitmeaastased rohttaimed, mõnikord üheaastased taimed. Üheaastaste taimede olemasolu peetakse evolutsiooniliselt nooreks tunnuseks. Lehtede asetus on vahelduv, lehed on lihtsad, ilma lehtedeta ja võivad olla terved või tükeldatud.
Mooniõied on sageli suured ja aktinomorfsed:
* Ca 2 Co 2 + 2 A ¥ G ( ¥ )
Reeglina kukuvad tupplehed õie avanedes maha. Günoetsium koosneb mitmest karpast, mis kasvavad kokku, moodustades parakarpse günoetsiumi. Vili on kuiv kapsel, mis on kaetud nelkidega. Munarakke on palju, viljad on väikesed ja sisaldavad varumaterjalina rasvu.
Mooni silmatorkav anatoomiline ja biokeemiline omadus on latiferi olemasolu, milles sünteesitakse keeruka struktuuriga piimjas mahl. See sisaldab erinevaid isokinoliini alkaloide. Mooniseemnete piimmahla ja alkaloide kasutatakse laialdaselt meditsiinis.
Mooniperekond on omandanud erilise tähtsuse unimaguna liigid, mis on oopiumi tootmise peamine tooraine. Oopium sisaldab rohkem kui 20 alkaloidi, sealhulgas morfiini, kodeiini, narkotiini ja papaveriini. Mõnel alkaloidil on võimas anesteetiline toime, kuid need tekitavad sõltuvust. Papaveriini kasutatakse spasmolüütikumina stenokardia ja bronhiaalastma korral. Meie maal uinutavat mooni ei kasvatata. Teatud koguses narkootilisi alkaloide sisaldavad ka õliseemned ja mooni dekoratiivsordid.
Suurepärane vereurmarohi
Lilled on väikesed, kogutud vihmakujulisse õisikusse: * Ca 2 Co 2 +2 A ¥ G (¥)
Selle apelsini piimjas mahl koos alkaloididega sisaldab flavonoide, tanniine, saponiine, orgaanilisi happeid ja vitamiine. Vereurmarohi kasutatakse kolereetilise ja antibakteriaalse vahendina maksa- ja sapipõiehaiguste korral. Suur vereurmarohi on mürgine taim.
Ravim "Sangviritrin" on valmistatud vereurmarohist suu ja kurgu loputamiseks, kui need on põletikulised, samuti mädaste haavade pesemiseks.
Kollast mooniseemneid kasutatakse meditsiinilistel eesmärkidel ja kasutatakse selle saamiseks Mitte narkootiline köhavastane glautsiinvesinikkloriid. Meditsiinilistel eesmärkidel kasutatakse ka väikeseviljalist makki ja südamikku.
28. ÕPPETUND: Rosaceae perekond.
1. Rosaceae perekond, üldtunnused.
2. Alamsugukond Spiraea. Alamperekond roosa.
3. Apple'i alamperekond. Ploomi alamperekond.
1. Rosaceae perekond (3500)
Selle perekonna esindajad on levinud väljaspool põhjapoolkera troopilisi piirkondi ja neid leidub erinevates fütotsenoosides. Smolenski piirkonnas kasvab 56 liiki Rosaceae. Eluvorm: puud, põõsad, maitsetaimed (ühe- ja mitmeaastased).
Lehed on nii lihtsad (terved või tükeldatud) kui ka komplekssed (paaritud ja ebaharilikud, kolmelehelised), väga sageli esineb lehtpuid. Lehtede paigutus on vahelduv, harva vastupidine. Mõnel liigil on ogad (epidermise metamorfoosid).
Rosaatseae biokeemilised omadused:
· puudub spetsialiseerimata sekundaarne ainevahetus. See väljendub alkaloidide ja toksiinide täielikus puudumises. Eeterlikud õlid moodustuvad harva ning neil on alati lihtne koostis ja struktuur (välja arvatud perekond Rose)
Rosaceae lillede struktuur ei ole eriti spetsialiseerunud putukatolmlemiseks. Lilled üksikult või kogutud õisikutesse erinevat tüüpi. Õied on aktinomorfsed, periant on kompleksne – tupp- ja kroonlehti on tavaliselt viis. Üsna sageli on lillel kahekordne tupplehtede arv, mille puhul moodustub esimene tupplehtede ring kehvem. Tolmukaid on palju, 2-4 korda rohkem kui kroonlehti. Karpeldade arv on kas määramata või rangelt fikseeritud.
Iseloomulik tunnus lill – kohalolek hüpantium- spetsiaalne moodustis, mis moodustub kinnikasvanud anumast ja sellega kokku sulanud tupplehtede, kroonlehtede ja tolmukate alustest. Hüpantiumi kuju võib olla kumer, taldrikukujuline või nõgus. Hüpantium kaitseb günoetsiumi keskkonnategurite eest. Samal ajal mängib see olulist rolli puuviljade moodustamisel. See võib kasvada ja osaleda mesokarpide ja endokarpide moodustamises. Seega on kibuvitsamarjade, õunapuude, pirnide ja ploomide mahlane viljaliha ülekasvanud hüpantium. Lisaks pakub hüpanthium sidet loomadega, kes jagavad puuvilju (ekso- ja endokooria).
Rosaceae viljad on struktuurilt väga mitmekesised, mis pakub erinevaid levitamisviise. Jaotusmeetodite täiustamine on perekonnasisese evolutsiooni üks peamisi suundi.
Väga sageli toodavad Rosaceae võltsvilju (õun õunapuus ja õun pihlakas) ja keerulisi vilju (polupähkel maasikates ja polüpähklid vaarikates).
Rosaceae perekonda kuulub neli alamperekonda: spireaceae, roseaceae, õun ja ploom.
2. Alamsugukond Spiraea. Alamperekond roosa.
Alamperekond Spiraea (180)
See on kõige primitiivsem alamperekond, mida esindavad põõsad, harvemini puud ja väga harva mitmeaastased risoomirohud. Lilled on väikesed, kogutud kas panicles, ratsemoosi õisikutesse või korümbiinidesse. Korolla kroonlehed on valged, harvem heleroosad. Selle perekonna esindajaid iseloomustab nõgus või peaaegu tasane anum (hüpantium) ja vili on kuiv - mitmeleheline (kvintaleheline). Günoetsium on apokarpne, koosneb tavaliselt 2-5 karpast.
Perekonda kuuluvad perekonnad Spirea, Meadowsweet ja Rowanberry.
Keskperekond on perekond Spiraea, levinud Siberis ja Kaug-Ida. Spiraea on lihtsate lehtedega põõsad, millel pole varre. Õied on tavaliselt valged või roosad, paniculate, corymbose või umbellate õisikutes. Vili on mitmeleheline.
Perekond nurmenukk, liik nurmenukk. See on suur, kuni 2 m kõrgune lühikese risoomiga mitmeaastane taim. Lehed on vahelduvalt sulgjad, ülalt tumerohelised, alt õhukese valge-tomentoosse karvaga. Hõõrumisel eritavad lehed teravat lõhna. Lilled on kollakasvalged ja kogutud tihedatesse paniculate õisikutesse. Viljad on üheseemnelised lehekesed. Meadowsweet on kasutatud pikka aega rahvameditsiin kokkutõmbajana.
* Ca (5) Co 5 A ¥ G 6-10
Alamsugukond Rosaceae (1700)
Rohttaimed levinud erinevates fütotsenoosides tundrast kuni mägise troopikani. Lilled kogutakse korümboosi või ratsemoosi dihhaasiasse, harvemini üksikuid lilli.
koerroos – * Ca (5) Co 5 A ¥ G ¥
Günoetsium on apokarpne, polümeerne, kuid munasarjas on alati üks, harva kaks munarakku.
Viljad on mitmekesised, kuid enamasti on need koondunud luuviljad ja mitmepähklid. Paljudel selle perekonna liikidel on mahlased komplekssed viljad, mille moodustamisel osaleb hüpantium.
Paljusid selle perekonna taimi iseloomustab paljunemine risoomide, stoolide või kõõlustega.
Rooside sekundaarse metabolismi aineid kasutatakse ravimitena.
1. harilik vaarikas - selle viljad sisaldavad märkimisväärses koguses salitsüülhapet, mis määrab nende diaforeetilise ja palavikualandaja toime külmetushaiguste vastu.
2. Burnet - õiel puudub korolla, tupp on lilla ja 4-osaline. Risoomi keedust kasutatakse hemostaatilise ainena emaka ja hemorroidide verejooksu korral.
3. Potentilla erecta (Uzik, Kalgan). Seda eristavad 4-liikmeline pärand ja alamkupuga tupp. Galangali tinktuuri (25g risoomi 500ml viina kohta) kasutatakse soolekatarri ja düsenteeria korral.
4. sookakk (rahvapärane nimetus - dekop) - risoomi keedust kasutatakse liigesehaiguste ja kollatõve puhul. Kasutatakse ka kokkutõmbava, hemostaatilise ja diaphoreetilise vahendina.
5. kaneeli kibuvits - sisaldab suures koguses C-vitamiini, samuti vitamiine B2, K, karoteeni ja sidrunhape. Kolereetilise ravimina kasutatakse kibuvitsa kondenseeritud vesiekstrakti suhkruga (holosas).
3. Apple'i alamperekond. Ploomi alamperekond.
 PUNAVETIKATE OSAKOND on peaaegu eranditult merevetikad. elutsükkel - lipufaaside täielik puudumine oogaamse seksuaalprotsessi erivorm; 2
PUNAVETIKATE OSAKOND on peaaegu eranditult merevetikad. elutsükkel - lipufaaside täielik puudumine oogaamse seksuaalprotsessi erivorm; 2
 Pigmendid klorofüll “a” ja “b”, karotenoidid (karoteen, zeaksantiin, anteraksantiin, krüptoksantiin, luteiin, neoksantiin) fükobiliinid: fükoerütriinid (punased) fükotsüaniinid ja allofükotsüaniin (sinine) terasest kuni terase värvusega tõlkes. fükotsüaniini liig) 3
Pigmendid klorofüll “a” ja “b”, karotenoidid (karoteen, zeaksantiin, anteraksantiin, krüptoksantiin, luteiin, neoksantiin) fükobiliinid: fükoerütriinid (punased) fükotsüaniinid ja allofükotsüaniin (sinine) terasest kuni terase värvusega tõlkes. fükotsüaniini liig) 3
 Kloroplastid on kahest membraanist, üksikutest tülakoididest koosnev kest fükobilisoomi pinnal. Varutoode on polüsahhariid “lilla tärklis”, mis omandab joodilt pruunikaspunase värvuse. Purpursed tärkliseterad ladestuvad tsütoplasmas alati ilma ühenduseta pürenoidide ja kromatofooridega. 4
Kloroplastid on kahest membraanist, üksikutest tülakoididest koosnev kest fükobilisoomi pinnal. Varutoode on polüsahhariid “lilla tärklis”, mis omandab joodilt pruunikaspunase värvuse. Purpursed tärkliseterad ladestuvad tsütoplasmas alati ilma ühenduseta pürenoidide ja kromatofooridega. 4
 Talluse struktuur on üherakulised kookoidvormid (porfüüridium), heterotrihholised ja hargnenud filamentide kujul, mis on risoidide abil substraadi külge kinnitatud. pseudoparenhümatoosne talli, külgharude põimumine Parenhümatoosse struktuuriga lamelltalli (porfüür). 5
Talluse struktuur on üherakulised kookoidvormid (porfüüridium), heterotrihholised ja hargnenud filamentide kujul, mis on risoidide abil substraadi külge kinnitatud. pseudoparenhümatoosne talli, külgharude põimumine Parenhümatoosse struktuuriga lamelltalli (porfüür). 5
 Rakk on kaetud pektiinkestaga – hemitselluloosi komponendid paisuvad ja seintesse ladestub lubi. Rakud on ühe- ja mitmetuumalised. Kromatofoorid on parietaalsed, arvukad, terade või plaatide kujul. 6
Rakk on kaetud pektiinkestaga – hemitselluloosi komponendid paisuvad ja seintesse ladestub lubi. Rakud on ühe- ja mitmetuumalised. Kromatofoorid on parietaalsed, arvukad, terade või plaatide kujul. 6
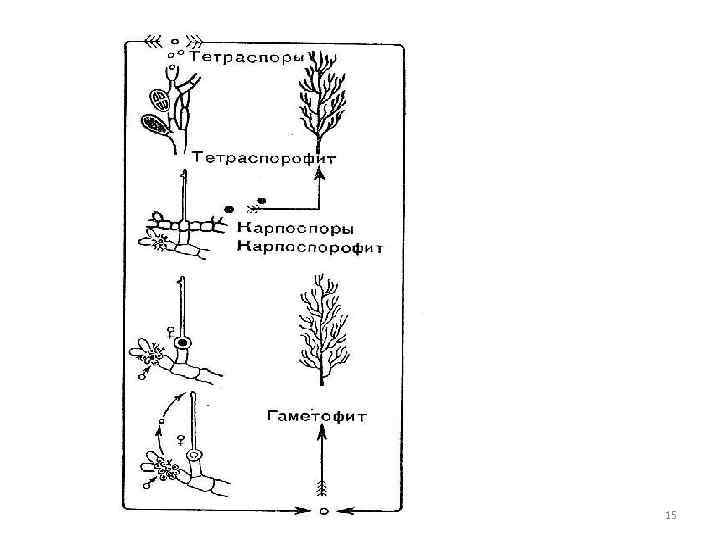
 Mittesuguline paljunemine liikumatute rakkude kaudu monospoorid - üks sporangiumis Tetraspoorid - neli tetraspoorit - diploidsetel aseksuaalsetel taimedel - sporofüüdid. Sporangiumides tekib meioos enne tetraspooride moodustumist. 7
Mittesuguline paljunemine liikumatute rakkude kaudu monospoorid - üks sporangiumis Tetraspoorid - neli tetraspoorit - diploidsetel aseksuaalsetel taimedel - sporofüüdid. Sporangiumides tekib meioos enne tetraspooride moodustumist. 7
 Seksuaalne protsess Oogamous Naisorgan- enamuses karpogon - laiendatud basaalosast - kõht - muna ja protsess - trikohonid. Carpogon areneb spetsiaalsel lühikesel karpogonaalsel oksal. 8
Seksuaalne protsess Oogamous Naisorgan- enamuses karpogon - laiendatud basaalosast - kõht - muna ja protsess - trikohonid. Carpogon areneb spetsiaalsel lühikesel karpogonaalsel oksal. 8
 Anteridia - väikesed värvitud rakud, mis sisaldavad lipustunud spermatosoide, spermatosoidid transporditakse passiivselt veevoolude toimel ja kleepuvad trikogiini külge. Kokkupuutekohas sperma ja trikhoogiinid lahustuvad nende seinad, spermatosoidide tuum liigub mööda trihhogiini karpogoni kõhuosas, ühineb 9-ga.
Anteridia - väikesed värvitud rakud, mis sisaldavad lipustunud spermatosoide, spermatosoidid transporditakse passiivselt veevoolude toimel ja kleepuvad trikogiini külge. Kokkupuutekohas sperma ja trikhoogiinid lahustuvad nende seinad, spermatosoidide tuum liigub mööda trihhogiini karpogoni kõhuosas, ühineb 9-ga.
 karpospooride moodustumine Pärast viljastamist eraldatakse karpogoni basaalosa vaheseinaga trikoosist - see sureb Areng - karpospooride teke sigooti sisu (viljastatud karpogon) 1. jaguneb otse, moodustades liikumatud paljad eosed - karpospoorid, 10
karpospooride moodustumine Pärast viljastamist eraldatakse karpogoni basaalosa vaheseinaga trikoosist - see sureb Areng - karpospooride teke sigooti sisu (viljastatud karpogon) 1. jaguneb otse, moodustades liikumatud paljad eosed - karpospoorid, 10
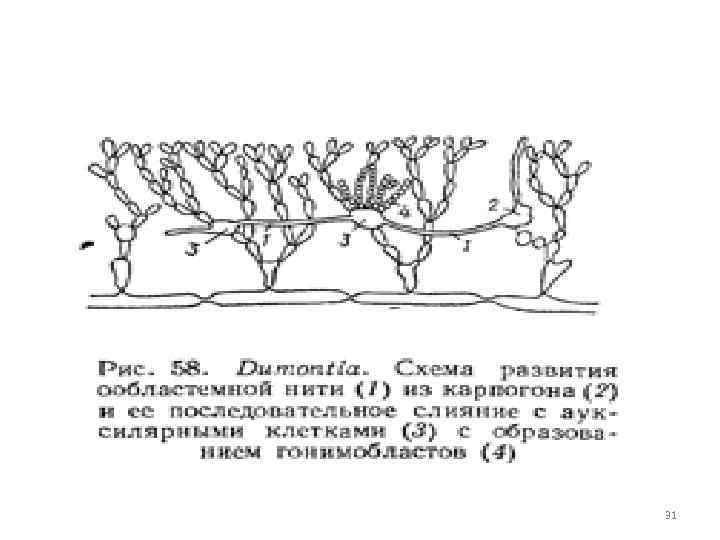
 karpospooride teke 2. viljastatud karpogonist kasvavad hargnevad niidid - gonimoblastid, nende rakud muutuvad karposporangideks, tekitades korraga ühe karpospoore. 11
karpospooride teke 2. viljastatud karpogonist kasvavad hargnevad niidid - gonimoblastid, nende rakud muutuvad karposporangideks, tekitades korraga ühe karpospoore. 11
 karpospooride moodustumine enamikus gonimoblastides ei arene otse aksoksülaarsetest rakkudest viljastatud karpogoni kõhust. saab eemaldada karpogonist või asuvad tallil vahetus läheduses, kui abirakud eemaldatakse karpogonist, selle kõhust kasvavad pärast viljastamist siderakud või oblastid y e, niidid. 12
karpospooride moodustumine enamikus gonimoblastides ei arene otse aksoksülaarsetest rakkudest viljastatud karpogoni kõhust. saab eemaldada karpogonist või asuvad tallil vahetus läheduses, kui abirakud eemaldatakse karpogonist, selle kõhust kasvavad pärast viljastamist siderakud või oblastid y e, niidid. 12
 oblastiliste filamentide rakud sisaldavad diploidseid tuumasid. Oblastilised niidid kasvavad abirakkude poole, kokkupuutepunktis membraanid lahustuvad ja tekib side oblastilise filamendi raku ja abiraku vahel. Selle rakkude liitumisega ei kaasne nende tuumade (ooblastilise filamendi diploidse rakutuuma ja abifilamendi haploidse tuuma) ühinemist. ühinemine abirakuga stimuleerib oblasti filamentraku diploidse tuuma jagunemist ja gonimoblastide arengut, mis sisaldavad diploidseid tuumasid ja toodavad diploidseid karpospoore Gonimoblastid - eripõlvkond - 13
oblastiliste filamentide rakud sisaldavad diploidseid tuumasid. Oblastilised niidid kasvavad abirakkude poole, kokkupuutepunktis membraanid lahustuvad ja tekib side oblastilise filamendi raku ja abiraku vahel. Selle rakkude liitumisega ei kaasne nende tuumade (ooblastilise filamendi diploidse rakutuuma ja abifilamendi haploidse tuuma) ühinemist. ühinemine abirakuga stimuleerib oblasti filamentraku diploidse tuuma jagunemist ja gonimoblastide arengut, mis sisaldavad diploidseid tuumasid ja toodavad diploidseid karpospoore Gonimoblastid - eripõlvkond - 13
 Kõige paremini organiseeritud punavetikates arenevad abirakud alles pärast karpogonumi viljastamist ja selle vahetus läheduses. Abiraku (või -rakkude) kombinatsiooni karpogoniga nimetatakse spetsiaalselt prokarpiks. Karpogoni ja abirakke ühendavate pikkade ooblastiliste filamentide moodustamine puudub vajadus, et abirakk lihtsalt sulandub viljastatud karpogoni kõhuga, mille järel arenevad sellest karpospooridega gonimoblastid - tsüstokarpid, kaetud pseudoparenhümatoosse membraaniga, mis areneb karpogoniga külgnevatest rakkudest. 14
Kõige paremini organiseeritud punavetikates arenevad abirakud alles pärast karpogonumi viljastamist ja selle vahetus läheduses. Abiraku (või -rakkude) kombinatsiooni karpogoniga nimetatakse spetsiaalselt prokarpiks. Karpogoni ja abirakke ühendavate pikkade ooblastiliste filamentide moodustamine puudub vajadus, et abirakk lihtsalt sulandub viljastatud karpogoni kõhuga, mille järel arenevad sellest karpospooridega gonimoblastid - tsüstokarpid, kaetud pseudoparenhümatoosse membraaniga, mis areneb karpogoniga külgnevatest rakkudest. 14
 15
15

 BANGUY KLASS Rakud, millel on sageli tähtkromatofoor ja pürenoid. Rakkudevahelised poorid tavaliselt puuduvad. Karpogon ilma trikogiinita jagatakse karpogoni sisu pärast viljastamist otse karpospooride moodustamiseks. Mittesuguline paljunemine – monospoorid 17
BANGUY KLASS Rakud, millel on sageli tähtkromatofoor ja pürenoid. Rakkudevahelised poorid tavaliselt puuduvad. Karpogon ilma trikogiinita jagatakse karpogoni sisu pärast viljastamist otse karpospooride moodustamiseks. Mittesuguline paljunemine – monospoorid 17

 19
19
 perekond compsopogon Magevesi troopikas laialt levinud koos akvaariumitaimedega Thallus heterotrichous Paljunemine monospooride abil. Monosporangiumid eraldatakse talluse mis tahes rakust kaldus vaheseinaga ja nende sisu on purpurse tärklise rohkusest tingitud granuleeritud sisu, mis muutub üheks liikumatuks paljaks monospooriks 20
perekond compsopogon Magevesi troopikas laialt levinud koos akvaariumitaimedega Thallus heterotrichous Paljunemine monospooride abil. Monosporangiumid eraldatakse talluse mis tahes rakust kaldus vaheseinaga ja nende sisu on purpurse tärklise rohkusest tingitud granuleeritud sisu, mis muutub üheks liikumatuks paljaks monospooriks 20
 21
21

 KLASS FLORIDA Rakud enamasti parietaalsete kromatofooridega ilma pürenoidideta Rakkude vahel on poorid. Carpogon trikogiiniga. Pärast viljastamist arenevad gonimoblastid kas otse viljastatud karpogoni kõhust või abirakkudest pärast nende sulandumist oblastiliste filamentidega. Enamiku osade mittesuguline paljunemine toimub tetraspooride abil. Karposporangiat kandvaid gonimoblaste peetakse eripõlvkonnaks – karposporofüüdiks. Karposporofüüdi arengu üksikasjad (viljastatud karpogonist või abirakkudest), samuti abirakkude diferentseerumise aeg (enne või pärast viljastamist), nende asend tallil jne moodustavad aluse jagunemisel. Floridia kuueks orduks 23
KLASS FLORIDA Rakud enamasti parietaalsete kromatofooridega ilma pürenoidideta Rakkude vahel on poorid. Carpogon trikogiiniga. Pärast viljastamist arenevad gonimoblastid kas otse viljastatud karpogoni kõhust või abirakkudest pärast nende sulandumist oblastiliste filamentidega. Enamiku osade mittesuguline paljunemine toimub tetraspooride abil. Karposporangiat kandvaid gonimoblaste peetakse eripõlvkonnaks – karposporofüüdiks. Karposporofüüdi arengu üksikasjad (viljastatud karpogonist või abirakkudest), samuti abirakkude diferentseerumise aeg (enne või pärast viljastamist), nende asend tallil jne moodustavad aluse jagunemisel. Floridia kuueks orduks 23
 Nemalionaceae seltsi esindajaid iseloomustab abirakkude puudumine gonimoblastid, mis arenevad otse viljastatud karpogonist 24
Nemalionaceae seltsi esindajaid iseloomustab abirakkude puudumine gonimoblastid, mis arenevad otse viljastatud karpogonist 24



 Perekonda Lemanea leidub kiirevoolulistes jõgedes koos külm vesi. Tallus on 10–15 cm pikkuste ja 1 mm paksuste mittehargnevate harjaste välimusega, tumelilla või oliivpruuni värvusega koos sõlmeliste tursetega. Kinnitatakse aluspinnale roomavatest niitidest valmistatud talla abil. Üks piklike värvitute rakkude niit jookseb piki talluse telge. Kesktelje iga lahtri tipust ulatub enamasti neljast kiirgavast harust koosnev keeris. Iga sellise haru basaalrakud on suured ja piklikud. Nende distaalsest (morfoloogiliselt ülemisest) otsast ulatuvad teist järku oksad, mis omakorda hargnevad korduvalt. Lõppoksad kasvavad kokku mitmekihiliseks kooreks. Selle välimised rakud on väikesed ja täidetud kromatofooridega, 28
Perekonda Lemanea leidub kiirevoolulistes jõgedes koos külm vesi. Tallus on 10–15 cm pikkuste ja 1 mm paksuste mittehargnevate harjaste välimusega, tumelilla või oliivpruuni värvusega koos sõlmeliste tursetega. Kinnitatakse aluspinnale roomavatest niitidest valmistatud talla abil. Üks piklike värvitute rakkude niit jookseb piki talluse telge. Kesktelje iga lahtri tipust ulatub enamasti neljast kiirgavast harust koosnev keeris. Iga sellise haru basaalrakud on suured ja piklikud. Nende distaalsest (morfoloogiliselt ülemisest) otsast ulatuvad teist järku oksad, mis omakorda hargnevad korduvalt. Lõppoksad kasvavad kokku mitmekihiliseks kooreks. Selle välimised rakud on väikesed ja täidetud kromatofooridega, 28
 Krüptoneemia järjekord sisaldab abirakke, mis arenevad enne karpogoni viljastamist ja on hajutatud kogu talluses teatud kaugusel karpogonist. Karpogonist kasvavad pärast viljastamist abirakkudeni enam-vähem pikad mitmerakulised side- ehk ooblastilised niidid. Pärast abiraku sulandumist ooblastilise filamendi rakuga ja filamentraku diploidse tuuma üleminekut abirakuks tekivad gonimoblastid, mis arenevad gonimoblastidel (karposporofüütidel), sisaldavad diploidset tuuma ja idanevad diploidseteks taimedeks. - tetrasporofüüdid, mis toodavad ainult mittesugulise paljunemise organeid - tetrasporangia. Tetraspooride moodustumisel tekib meioos ja haploidsed tetraspoorid idanevad haploidseteks gametofüüttaimedeks, mis kannavad paljunemisorganeid. Gametofüüt ja tetrasporofüüt ei erine väliselt (morfoloogiliselt). isomorfne põlvkondade vahetus, mida komplitseerib diploidne karposporoftoos. 29
Krüptoneemia järjekord sisaldab abirakke, mis arenevad enne karpogoni viljastamist ja on hajutatud kogu talluses teatud kaugusel karpogonist. Karpogonist kasvavad pärast viljastamist abirakkudeni enam-vähem pikad mitmerakulised side- ehk ooblastilised niidid. Pärast abiraku sulandumist ooblastilise filamendi rakuga ja filamentraku diploidse tuuma üleminekut abirakuks tekivad gonimoblastid, mis arenevad gonimoblastidel (karposporofüütidel), sisaldavad diploidset tuuma ja idanevad diploidseteks taimedeks. - tetrasporofüüdid, mis toodavad ainult mittesugulise paljunemise organeid - tetrasporangia. Tetraspooride moodustumisel tekib meioos ja haploidsed tetraspoorid idanevad haploidseteks gametofüüttaimedeks, mis kannavad paljunemisorganeid. Gametofüüt ja tetrasporofüüt ei erine väliselt (morfoloogiliselt). isomorfne põlvkondade vahetus, mida komplitseerib diploidne karposporoftoos. 29
 Lõunameredes levinud perekond Durenea. Tallus näeb välja nagu hargnenud, limased, roosad põõsad. 30
Lõunameredes levinud perekond Durenea. Tallus näeb välja nagu hargnenud, limased, roosad põõsad. 30
 31
31
 32
32
 33
33


 Liigid Ceramiaceae on kõrgelt organiseeritud, paljud abirakkude liigid diferentseeruvad alles pärast seda, kui on toimunud karpogoni viljastumine: karpogoni kõht sulandub kas vahetult külgneva(te) abiraku(te)ga. , moodustades tsüstokarpe 36
Liigid Ceramiaceae on kõrgelt organiseeritud, paljud abirakkude liigid diferentseeruvad alles pärast seda, kui on toimunud karpogoni viljastumine: karpogoni kõht sulandub kas vahetult külgneva(te) abiraku(te)ga. , moodustades tsüstokarpe 36


 Banguiaceae osakonna areng on primitiivsem. Carpogon ei ole veel välja kujunenud tüüpilist vormi ja erineb vähe tavalistest vegetatiivsetest rakkudest. Pärast viljastamist jagatakse karpogoni sisu otse karpospoorideks. Floridai on rohkem arenenud Carpogoni rühm koos Trichogynaga. Floridia klassist on lihtsaim järg nemalionaceae, millel puuduvad abirakud, ja gonimoblastid, millel arenevad karposporangiumid, moodustuvad otse viljastatud karpogoni kõhuosast. Krüptoneemia on järkjärgulise evolutsiooni järgmine samm: neil on abirakud, mis aitavad kaasa karpospooride tootmise suurenemisele, kuna moodustub mitte üks tsüstokarp, vaid palju - vastavalt abirakkude arvule. Kui abirakud on juhuslikult kogu talluses hajutatud, on vaja enam-vähem pikki oblastilisi filamente. Evolutsiooni kõrgeima faasi on jõudnud keramiidid, millel on prokarp ja abirakud diferentseeruvad alles pärast viljastumist. Karpogoni ja abiraku(de) lähedus prokarpis hõlbustab cpstokarpide teket. See järjekord on liigirikkaim. 39
Banguiaceae osakonna areng on primitiivsem. Carpogon ei ole veel välja kujunenud tüüpilist vormi ja erineb vähe tavalistest vegetatiivsetest rakkudest. Pärast viljastamist jagatakse karpogoni sisu otse karpospoorideks. Floridai on rohkem arenenud Carpogoni rühm koos Trichogynaga. Floridia klassist on lihtsaim järg nemalionaceae, millel puuduvad abirakud, ja gonimoblastid, millel arenevad karposporangiumid, moodustuvad otse viljastatud karpogoni kõhuosast. Krüptoneemia on järkjärgulise evolutsiooni järgmine samm: neil on abirakud, mis aitavad kaasa karpospooride tootmise suurenemisele, kuna moodustub mitte üks tsüstokarp, vaid palju - vastavalt abirakkude arvule. Kui abirakud on juhuslikult kogu talluses hajutatud, on vaja enam-vähem pikki oblastilisi filamente. Evolutsiooni kõrgeima faasi on jõudnud keramiidid, millel on prokarp ja abirakud diferentseeruvad alles pärast viljastumist. Karpogoni ja abiraku(de) lähedus prokarpis hõlbustab cpstokarpide teket. See järjekord on liigirikkaim. 39
 Vanim punavetikana identifitseeritud fossiil on ka vanim eukarüootne fossiil, mis kuulub konkreetsesse tänapäeva taksonisse. Arktilisest Kanadast leitud mitmerakuline fossiil Bangiomorpha pubescens erineb tänapäevasest punavetikate perekonnast Bangia vaid veidi, hoolimata sellest, et see ladestus kihtidena 1,2 miljardit aastat tagasi. 41
Vanim punavetikana identifitseeritud fossiil on ka vanim eukarüootne fossiil, mis kuulub konkreetsesse tänapäeva taksonisse. Arktilisest Kanadast leitud mitmerakuline fossiil Bangiomorpha pubescens erineb tänapäevasest punavetikate perekonnast Bangia vaid veidi, hoolimata sellest, et see ladestus kihtidena 1,2 miljardit aastat tagasi. 41
 DIVISION BROWN AGAE RNAEORNUTH mereline, eriti rohkesti põhja- ja lõunapoolkera külmades vetes. Vastavalt talluse morfoloogilisele ja anatoomilisele diferentseerumisele - rohkem kõrgel tasemel kui varem käsitletud vetikate rühmad. Ei ole teada ei ainurakseid ega koloniaalvorme ega talli lihtsa hargnemata niidi kujul. kõige lihtsamad on heterotrioossed tallid, suured, vale või tõelise koestruktuuriga.
DIVISION BROWN AGAE RNAEORNUTH mereline, eriti rohkesti põhja- ja lõunapoolkera külmades vetes. Vastavalt talluse morfoloogilisele ja anatoomilisele diferentseerumisele - rohkem kõrgel tasemel kui varem käsitletud vetikate rühmad. Ei ole teada ei ainurakseid ega koloniaalvorme ega talli lihtsa hargnemata niidi kujul. kõige lihtsamad on heterotrioossed tallid, suured, vale või tõelise koestruktuuriga.
 tugevalt limased rakuseinad, üks tuum, üks või mitu vakuooli, tavaliselt erineva kujuga seinakromatofoorid. kromatofoore ümbritseb keeruline membraanide süsteem - otseses ühenduses tuumaümbrisega - "kloroplasti endoplasmaatiline retikulum" Kloroplasti maatriksiga lõikuvad paralleelsed tritülakoidlamellid. Tülakoidivaba pürenoid ulatub kloroplastist välja neeru kujul.
tugevalt limased rakuseinad, üks tuum, üks või mitu vakuooli, tavaliselt erineva kujuga seinakromatofoorid. kromatofoore ümbritseb keeruline membraanide süsteem - otseses ühenduses tuumaümbrisega - "kloroplasti endoplasmaatiline retikulum" Kloroplasti maatriksiga lõikuvad paralleelsed tritülakoidlamellid. Tülakoidivaba pürenoid ulatub kloroplastist välja neeru kujul.
 Kromatofoori pigmendid on pruuni värvi, klorofüllid "a" ja "c" (klorofüll "b" puuduvad) β-karoteen Pruuni ksantofülle on palju, eriti fukoksantiini. 44
Kromatofoori pigmendid on pruuni värvi, klorofüllid "a" ja "c" (klorofüll "b" puuduvad) β-karoteen Pruuni ksantofülle on palju, eriti fukoksantiini. 44
 Varupolüsahhariid – laminariinalkohol mannitoolrasvad – ladestub tsütoplasmas väljapoole kloroplasti. Monaadirakkudel (zoospooridel ja sugurakkudel) on silm ja lipukesed. Ocellus on osa plastiidist ja on seotud lipuaparaadiga. Lipud on heteromorfsed. 45
Varupolüsahhariid – laminariinalkohol mannitoolrasvad – ladestub tsütoplasmas väljapoole kloroplasti. Monaadirakkudel (zoospooridel ja sugurakkudel) on silm ja lipukesed. Ocellus on osa plastiidist ja on seotud lipuaparaadiga. Lipud on heteromorfsed. 45


 Paljunemine on vegetatiivne, aseksuaalne ja seksuaalne. Vegetatiivne paljundamine talluse osade kaupa. Mõnel on spetsiaalsed oksad (haudmepungad), mis murduvad uuteks tallideks. Mittesuguline paljunemine - zoospoorid moodustuvad diploidsetel taimedel (sporofüüdid) ühekohalistes või ühekambrilistes sporangiumides, tuum jaguneb vähehaaval haploidseteks sugutaimedeks - gametofüütideks, millel moodustuvad suguelundid. Algloomadel arenevad sugurakud isogaamsetes või mitmekambrilistes sporangiumides. Oogoonias ja anteridiaas moodustub reeglina üks sugurakk (vastavalt munarakk ja seemnerakk). Munarakk viljastatakse alati väljaspool oogiumi. Sügoot kasvab diploidseks taimeks ilma puhkeperioodita.
Paljunemine on vegetatiivne, aseksuaalne ja seksuaalne. Vegetatiivne paljundamine talluse osade kaupa. Mõnel on spetsiaalsed oksad (haudmepungad), mis murduvad uuteks tallideks. Mittesuguline paljunemine - zoospoorid moodustuvad diploidsetel taimedel (sporofüüdid) ühekohalistes või ühekambrilistes sporangiumides, tuum jaguneb vähehaaval haploidseteks sugutaimedeks - gametofüütideks, millel moodustuvad suguelundid. Algloomadel arenevad sugurakud isogaamsetes või mitmekambrilistes sporangiumides. Oogoonias ja anteridiaas moodustub reeglina üks sugurakk (vastavalt munarakk ja seemnerakk). Munarakk viljastatakse alati väljaspool oogiumi. Sügoot kasvab diploidseks taimeks ilma puhkeperioodita.

 KLASS ISOGENERAATID Tellige Ectocarpous perekond Ectocarpus - kõige primitiivsemad pruunvetikad. Nad on levinud kõigis meredes, eriti külmades, ja kasvavad veealustel objektidel ja muudel suurematel vetikatel. 50
KLASS ISOGENERAATID Tellige Ectocarpous perekond Ectocarpus - kõige primitiivsemad pruunvetikad. Nad on levinud kõigis meredes, eriti külmades, ja kasvavad veealustel objektidel ja muudel suurematel vetikatel. 50
 51
51
 52
52

 54
54
 Selgkond Cutleriaceae Seltskonda kuulub ainult kaks perekonda: Cutleria ja Zanardinia Cutleria on levinud kogu Euroopa rannikul, Zanardinia - peamiselt Vahemeres, aga ka Mustas meres. Mõlemal perekonnal on põlvkondade vaheldumine: Cutlerias on põlvkondade vahetus heteromorfne, Zanardiinias isomorfne 55
Selgkond Cutleriaceae Seltskonda kuulub ainult kaks perekonda: Cutleria ja Zanardinia Cutleria on levinud kogu Euroopa rannikul, Zanardinia - peamiselt Vahemeres, aga ka Mustas meres. Mõlemal perekonnal on põlvkondade vaheldumine: Cutlerias on põlvkondade vahetus heteromorfne, Zanardiinias isomorfne 55

 57
57
 58
58

 60
60

 62
62
 KLASS HETEROGENERAATNE Laminaria Gametofüüdid erinevad üksteisest vähe ja neid esindavad mikroskoopilised, sageli mitmeks rakuks taandunud niitjad taimekesed, mis kannavad paljunemisorganeid. Isastel gametofüütidel moodustuvad anteridiad väikeste rakkude kujul, millest areneb üks sperma, emastel gametofüütidel - oogoonia, milles moodustub üks munarakk. Erinevate perekondade sporofüüdid erinevad järsult ja esindavad suurimaid madalamaid taimi, ulatudes 60–100 m pikkuseni, olulise morfoloogilise jagunemise ja kompleksiga. anatoomiline struktuur. Enamasti jagunevad pruunvetika sporofüüdid lehekujuliseks teraks, “tüveks” ja risoidideks, mille abil kinnitub kogu taim veealuste kivide ja kivide külge. Leheraba ja varre liitumiskohas on interkalaarne meristeem, mille tegevuse tõttu kasvavad nii lehelaba kui ka vars. pruunvetika perekond, mille liigid on laialt levinud põhjameres. 63
KLASS HETEROGENERAATNE Laminaria Gametofüüdid erinevad üksteisest vähe ja neid esindavad mikroskoopilised, sageli mitmeks rakuks taandunud niitjad taimekesed, mis kannavad paljunemisorganeid. Isastel gametofüütidel moodustuvad anteridiad väikeste rakkude kujul, millest areneb üks sperma, emastel gametofüütidel - oogoonia, milles moodustub üks munarakk. Erinevate perekondade sporofüüdid erinevad järsult ja esindavad suurimaid madalamaid taimi, ulatudes 60–100 m pikkuseni, olulise morfoloogilise jagunemise ja kompleksiga. anatoomiline struktuur. Enamasti jagunevad pruunvetika sporofüüdid lehekujuliseks teraks, “tüveks” ja risoidideks, mille abil kinnitub kogu taim veealuste kivide ja kivide külge. Leheraba ja varre liitumiskohas on interkalaarne meristeem, mille tegevuse tõttu kasvavad nii lehelaba kui ka vars. pruunvetika perekond, mille liigid on laialt levinud põhjameres. 63
 KLASS CYCLOSPORONAE Järjekorrale fucus on iseloomulik - - aseksuaalse paljunemise puudumine ogaamilise seksuaalse protsessi kaudu mõnes ookeani osas kuni tohutute Sargassum thalli kogunemiseni (näiteks Sargasso meri).
KLASS CYCLOSPORONAE Järjekorrale fucus on iseloomulik - - aseksuaalse paljunemise puudumine ogaamilise seksuaalse protsessi kaudu mõnes ookeani osas kuni tohutute Sargassum thalli kogunemiseni (näiteks Sargasso meri).
 perekond Sargassum talluse kompleksne morfoloogiline jagunemine on levinud lõunapoolkeral, eriti soojades meredes. Vars kinnitub aluspinnale tallaga, okstega, kannab lapikuid lehelaadseid moodustisi, kerakujulisi õhumulle spetsiaalsetel vartel, harunenud viljakaid oksi 71
perekond Sargassum talluse kompleksne morfoloogiline jagunemine on levinud lõunapoolkeral, eriti soojades meredes. Vars kinnitub aluspinnale tallaga, okstega, kannab lapikuid lehelaadseid moodustisi, kerakujulisi õhumulle spetsiaalsetel vartel, harunenud viljakaid oksi 71



 Pruunvetikad on looduslik rühm, mis oma morfoloogias eristub hästi teistest vetikaliikidest. Kuid nagu juba mainitud, vastavalt mitmetele omadustele (klorofüllide "a" ja "c" olemasolu, klorofülli "b" puudumine, tülakoidide asukoht kloroplastides - tritüülakoidlamellid, sarnaste säilitustoodete järgi - pruunvetikas , krüsolamineriin, vastavalt monaadirakkude struktuurile heterokont ja heteromorfsete lipudega) pruunvetikatel on sarnasusi kuldsete, kollakasroheliste, ränivetikate ja pürofüütsete vetikatega. Selle põhjal taandavad mõned autorid pruunvetikate ja muud loetletud vetikate rühmad Credoshore'i ühe suure jaotuse klassidesse. autor 1 a. Monaadirakkude olemasolu pruunvetikate elutsüklis võimaldab neid, nagu ka teisi loetletud rühmi, tuletada mõnest primaarsest fotosünteetilisest flagellaadist, milles on ülekaalus pruunid pigmendid. Need flagellaadid arenesid mitmel viisil, millest üks viis pruunvetikate tekkeni. Siiski on pruunvetikatel lipudega kaugem seos kui kuldvetikatel, heteroflagellaatidel, ränivetikatel, pürofüütidel ja rohevetikatel, mis on mitmete omaduste poolest sarnased, aga ka rohevetikatel, kuna siin puuduvad otsesed üleminekuvormid. Pruunvetikate hulgas ei toimu talluse organisatsiooni tüüpide järjepidevat arengut monaadilisest niit- ja lamellvetikani, mida on lihtne täheldada roheliste, kollakasroheliste, kuldsete ja pürofüütsete vetikate puhul. Nagu juba märgitud, on pruunvetikate hulgas esindatud ainult talluse morfoloogilise diferentseerumise kõrgeim staadium - heterotrihaalne ja lamell. Võib-olla kadusid lihtsamad vormid (monaad, kookoid, lihtniitjad) pruunvetikate - juba tuntud iidse rühma - pika evolutsiooni käigus täielikult; Siluri ja Devoni leiukohtadest. Pruunvetikate osakonnasiseste liikide omavaheliste seoste ja nende asukoha kohta süsteemis ühest seisukohta ei ole. Vastavalt ühele pruunvetikate fülogeneesi skeemidest, mis on ehitatud maksimaalselt talluse struktuuri sarnasust arvesse võttes. varajased staadiumid areng, pruunvetikad - difüleetiline rühm: üks evolutsiooniliin ühendab (esitluses nimetatutest) Sphacelariaceae, Dictyoteceae ja Fucus, teine - Ectocarpaceae, Cutleriaceae ja Laminariaceae. Mõlemad 219" 75
Pruunvetikad on looduslik rühm, mis oma morfoloogias eristub hästi teistest vetikaliikidest. Kuid nagu juba mainitud, vastavalt mitmetele omadustele (klorofüllide "a" ja "c" olemasolu, klorofülli "b" puudumine, tülakoidide asukoht kloroplastides - tritüülakoidlamellid, sarnaste säilitustoodete järgi - pruunvetikas , krüsolamineriin, vastavalt monaadirakkude struktuurile heterokont ja heteromorfsete lipudega) pruunvetikatel on sarnasusi kuldsete, kollakasroheliste, ränivetikate ja pürofüütsete vetikatega. Selle põhjal taandavad mõned autorid pruunvetikate ja muud loetletud vetikate rühmad Credoshore'i ühe suure jaotuse klassidesse. autor 1 a. Monaadirakkude olemasolu pruunvetikate elutsüklis võimaldab neid, nagu ka teisi loetletud rühmi, tuletada mõnest primaarsest fotosünteetilisest flagellaadist, milles on ülekaalus pruunid pigmendid. Need flagellaadid arenesid mitmel viisil, millest üks viis pruunvetikate tekkeni. Siiski on pruunvetikatel lipudega kaugem seos kui kuldvetikatel, heteroflagellaatidel, ränivetikatel, pürofüütidel ja rohevetikatel, mis on mitmete omaduste poolest sarnased, aga ka rohevetikatel, kuna siin puuduvad otsesed üleminekuvormid. Pruunvetikate hulgas ei toimu talluse organisatsiooni tüüpide järjepidevat arengut monaadilisest niit- ja lamellvetikani, mida on lihtne täheldada roheliste, kollakasroheliste, kuldsete ja pürofüütsete vetikate puhul. Nagu juba märgitud, on pruunvetikate hulgas esindatud ainult talluse morfoloogilise diferentseerumise kõrgeim staadium - heterotrihaalne ja lamell. Võib-olla kadusid lihtsamad vormid (monaad, kookoid, lihtniitjad) pruunvetikate - juba tuntud iidse rühma - pika evolutsiooni käigus täielikult; Siluri ja Devoni leiukohtadest. Pruunvetikate osakonnasiseste liikide omavaheliste seoste ja nende asukoha kohta süsteemis ühest seisukohta ei ole. Vastavalt ühele pruunvetikate fülogeneesi skeemidest, mis on ehitatud maksimaalselt talluse struktuuri sarnasust arvesse võttes. varajased staadiumid areng, pruunvetikad - difüleetiline rühm: üks evolutsiooniliin ühendab (esitluses nimetatutest) Sphacelariaceae, Dictyoteceae ja Fucus, teine - Ectocarpaceae, Cutleriaceae ja Laminariaceae. Mõlemad 219" 75
 Järgude rühmad erinevad ka talli kasvuviisi poolest: esimest iseloomustab tipukasv, teist interkalaarne kasv. Kilin (N. Ku 1sh), mille süsteem on praegusel ajal enim levinud, tuletab pruunvetikate ühisest esivanemast kolm evolutsiooniliini, mida ta esitab isogeneratiivsete, heterogeneratiivsete ja tsüklospoorsete klassidena; need põhinevad vetikate elutsüklite ja morfoloogilise struktuuri erinevustel. Suuri pruunvetikaid kasutatakse laialdaselt inimeste põllumajanduses. Nendest ekstraheeritakse algiini - liimiainet, mida kasutatakse tekstiili-, toiduainetööstuses ja mitmetes muudes tööstusharudes. Rannikumaades kasutatakse kaaliumi- ja lämmastikurikkaid merevetikate heitmeid väetisena ja ka loomasöödana. Mõned, näiteks pruunvetikas (merevetikad), on söödavad. 76
Järgude rühmad erinevad ka talli kasvuviisi poolest: esimest iseloomustab tipukasv, teist interkalaarne kasv. Kilin (N. Ku 1sh), mille süsteem on praegusel ajal enim levinud, tuletab pruunvetikate ühisest esivanemast kolm evolutsiooniliini, mida ta esitab isogeneratiivsete, heterogeneratiivsete ja tsüklospoorsete klassidena; need põhinevad vetikate elutsüklite ja morfoloogilise struktuuri erinevustel. Suuri pruunvetikaid kasutatakse laialdaselt inimeste põllumajanduses. Nendest ekstraheeritakse algiini - liimiainet, mida kasutatakse tekstiili-, toiduainetööstuses ja mitmetes muudes tööstusharudes. Rannikumaades kasutatakse kaaliumi- ja lämmastikurikkaid merevetikate heitmeid väetisena ja ka loomasöödana. Mõned, näiteks pruunvetikas (merevetikad), on söödavad. 76
Osakonna nimi tuleb kreekakeelsest sõnast rodoon(“Rodon”) – roosa. Punavetikate värvus on tingitud erinevatest pigmentide kombinatsioonidest. See ulatub hallist ja lillast kuni peaaegu mustani ning hõlmab ka kõiki punase ja roosa toone. Tugevalt valgustatud aladel leiduvad punased vetikad on kollase, pruuni või musta värvi, kuna nende rakkudes on palju valgust kaitsvaid karotenoide. Ühendab üherakulisi, koloniaal- ja mitmerakulisi organisme, mille kehaehitus on kookoidne, filamentne, pseudoparenhümatoosne ja parenhümatoosne. Kõiki esindajaid iseloomustab lipufaaside täielik puudumine elutsüklis. Talluse kuju on mitmekesine: niitjas, põõsas, koorikutaoline, lamelljas, vesikulaarne, kotitaoline jne. Elavad peamiselt meredes ja ookeanides (tavaliselt kinnitunud vormid), harvemini leidub neid magevees. Teada on umbes 4 tuhat liiki.
Raku struktuur. Punavetikarakk on eukarüootne: see on moodustanud oma membraanidega organellid: tuum, mitokondrid, plastiidid, Golgi aparaat jt. Punavetikate rakkudes on plastiide üks kuni mitu. Rakkude vahel on pooride ühendused. Lisaks välisele rakumembraanile on Rhodophyte rakkudel eriline moodustis – rakusein.
Enamasti on üks tuum, kuid on ka mitmetuumalisi esindajaid. Punavetika tuumad on väikesed. Mõne liigi puhul on teada endoreduplikatsioon ehk tuumagenoomi replikatsioon ilma mitoosita. See põhjustab polüploidsust või kromosoomide arvu mitmekordset suurenemist rakkudes. Mitokondrid lamedate kristallidega.
Punavetikate kloroplastid on erineva kujuga, tavaliselt paiknevad need piki rakuseinu või parietaalselt. Need plastiidid on enamasti ketta- ja lindikujulised. Kloroplastide kuju võib sõltuvalt vetikate vanusest muutuda. Iga kloroplast on ümbritsetud oma topeltmembraaniga ja kloroplasti endoplasmaatilist retikulumit ei ole. Kloroplastides olevad tülakoidid on üksikud, ei kogune rühmadesse ja asuvad üksteisest samal kaugusel. Üks või kaks tülakoidi paiknevad tavaliselt piki kloroplasti perifeeriat, paralleelselt selle membraaniga. Kloroplasti DNA esineb väikeste nukleoidide kujul, mis on hajutatud kogu kloroplasti stroomas. Iga nukleoid sisaldab mitut ringikujulist kloroplasti DNA molekuli.
Klorofüllidest on punavetikate plastiidides ainult klorofüll A, mida varjavad täiendavad pigmendid - fükobiliinid: punane fükoerütriin, sinine fükotsüaniin ja allofükotsüaniin. Need fükobiliinid paiknevad tülakoidide pinnal spetsiaalsetes moodustistes - poolkera- ja poolkettakujulistes fükobilisoomides.
Rakusein koosneb struktuursest fibrillaarsest fraktsioonist ja amorfsest maatriksist. Rakuseina tugevuse annavad tselluloosfibrillid, mis punavetikates moodustavad ebakorrapärase võrgustiku. Polüsahhariidi amorfse maatriksi olulisemad ained on agar, agaroidid ja karrageen. Need ained sünteesitakse Golgi aparaadi tsisternides, transporditakse seejärel raku pinnale ja kinnistatakse seina. Need moodustavad kuni 70% rakuseina kuivkaalust. Paljudel punavetikatel võib rakuseina peal olla küünenahk, mis koosneb peamiselt valkudest. Purpursete taimede hulgas on lubjastunud kestadega liike: neisse ladestub kaltsiit või aragoniit. Osa tallust või kogu tallust võib inkrusteerida.
Üldised omadused.
Peaaegu kõik punavetikad on fototroofid ja ehitavad oma keha fotosünteesi abil. Fotosünteesi produktideks on spetsiaalne lillakas tärklis, mis ladestub tsütoplasmas, mitte aga kloroplastis, nagu rohevetikates. Purpurne tärklis annab joodiga erkpunase värvuse. Oluline varuprodukt on madala molekulmassiga süsivesiniku floridosiid. Selle sisaldus mõne esindaja tallis võib ületada 10% kuivkaalust. See täidab osmoregulatoorset funktsiooni. Lisaks punavetikatele leidub floridosiidi sinivetikates ja krüptomonaadides. Selle kontsentratsioon rakkudes suureneb koos keskkonna soolsuse suurenemisega. Mõned punased taimed võivad säilitada ka mitmehüdroksüülseid alkohole.
Enamik punavetikaliike on mitmerakulised keerukad organismid, mille suurus võib ulatuda 1–2 meetrini ja ainult algelised esindajad on üherakulise või koloniaalse struktuuriga (joon. 17). Esineb nii ühe- kui ka mitmeaastaseid liike, tavaliselt 3–6-aastaseid. Lillade kehakuju on väga
Riis. 17 Välimus punavetikad: A – üherakulised vetikad Porphyridium; B – mitmerakuline vetikas Delesseria
mitmekesine. See võib olla: niitjas (karvataoline või jäme), lamelljas, tahke või kompleksselt lahtikasvanud väljakasvudega piki serva, silindriline, kortikaalne (koorikud, aluspinnale surutud kiled), korallikujuline. Punavetikate väliste vormide mitmekesisus tuleneb talluse mitmest eristumise tüübist: kookoidne, filamentne, heterofilamentne, pseudotide ja kangas. Rhodophyte tallid on kinnitatud risoidide või taldade kaudu.
Floridaaceae vetikate tallid on kõige keerulisemad. Nende tallidel on märke kudede diferentseerumisest koos rakkude spetsialiseerumisega. Nende talluses võib eristada: ajukoort, mis koosneb mitmest intensiivselt värvitud rakkude kihist; tuum, mis koosneb värvitutest rakkudest, mis on sageli kogutud niitidesse. Südamik ei täida mitte ainult transpordifunktsiooni, vaid ka mehaanilist funktsiooni, kuna see sisaldab paksude pikisuunaliste seintega niite. Paljude punavetikate koore ja südamiku vahel võib olla suurte värvitute rakkude vahekiht. Talluse kasv on kõige sagedamini interkalaarne (interkalaarne) ja apikaalne (apikaalne), harvem basaal.
Paljundamine.
Punastel vetikatel on kolm paljunemisviisi: vegetatiivne, mittesuguline ja seksuaalne.
Mittesuguline paljunemine läbi erinevate eoste abil. Eosed võivad paikneda ühes, kahes või neljas sporangiumis; neid nimetatakse vastavalt monospoorideks, bispoorideks ja tetraspoorideks. Tetraspoorid võivad paikneda tetrasporangiumides erineval viisil: üksteise kohal – lineaarselt ehk tsooniliselt, risti ja tetraeedri nurkades (joon. 18).

Riis. 18. Tetraspooride paigutuse tüübid tetrasporangiates (vastavalt: S. Noek van den et al., 1995): A– ristikujuline; B- ristikujuline pöörlemisega; IN- lineaarne; G – tetraeedriline
Seksuaalne paljunemine punavetikates on see oogaamne, tal on mitmeid tunnuseid, mida teistes vetikarühmades ei leidu. Meeste sugurakud - spermastumine, millel puuduvad flagellad ja nad kanduvad passiivselt veevooluga naiste suguelunditesse - karpogonid. Punaste vetikate karpogon on kolvi kujuga ja koosneb laiendatud alumisest osast (kõhust) ja piklikust ülemisest osast - trikohüüdid, mis on mõeldud spermatosoidide püüdmiseks. Enamikus punavetikates moodustub karpogon lühikese oksa, mida nimetatakse karpogonaalseks oksaks, otsa. Rakku, millest tekib karpogonaalne haru, nimetatakse toetavaks.
Spermatangiates tekivad ükshaaval spermatosoidid, mis omakorda moodustuvad emarakkudel. Küpsed spermatosoidid on mononukleaarsed ja neil puudub jäik rakusein, neid ümbritseb lima ja need võivad sisaldada kloroplaste. Sperma kandub passiivselt veevoolude toimel ja puutub kokku trikogiiniga, mis asub emasgametofüüdi pinna kohal. Spermatosoidide ja trikogiini seinad lahustuvad kokkupuutepunktis, isaste tuum läbib trikogoonis oleva tsentraalse kanali ja ühineb karpogooni haploidse tuumaga. Edasine areng erineva järgu liikidel on oma omadused.
Elutsükkel.
Enamikus Rhodophyta's ilmub pärast emassugurakkude - isassugurakkudega karpogonide - viljastamist sügoodist pärit spermatosoidid pärast mitme raku sulandumist mitmerakuline filamentne-parenhümaalne moodustis - karposporofüüt (joonis 19). Ta esindab

Riis. 19. Elutsükkel Batrachospermuma(pärast: R. E. Lee, 1999)
ise gonimoblast, milles arenevad diploidsed rakud – sugulise paljunemise eosed ehk karpospoorid, millest idaneb uus diploidne taim – sporofüüt. gonimoblast, ehk tsüstokarp on platsenta kompleks, millest ulatuvad välja gonimoblasti filamendid, mille otstes on karposporangiumid ja gonimoblastide kattekiht. Gonimoblastides on kõigil rakkudel diploidne kromosoomide komplekt.
Paljudes Floridaidae'is on gonimoblast ümbritsetud membraaniga. Sel juhul nimetatakse seda struktuuri tsüstokarpiks (joonis 20). Karposporangias

Riis. 20. Elutsükkel Polüsüfoonia(pärast: R. E. Lee, 1999)
moodustuvad karpospoorid, mis idanevad diploidseks tetrasporofüüdiks. Tetrasporofüüdil toimub redutseeriv jagunemine tetrasporangias. Haploidsed tetraspoorid idanevad haploidseks gametofüüdiks. See elutsükkel vahelduvate põlvkondadega: üks haploidne – gametofüüt ja kaks diploidset – karposporofüüt ja tetrasporofüüt – leidub enamikus punavetikates, kuid sellest on mitmeid kõrvalekaldeid, olenevalt tingimuste muutlikkusest nende keskkonnas.
Elutsükkel ei ole alati jäigalt fikseeritud. Kui ühe faasi arenguks tingimused puuduvad, siis ikka ja jälle, kuni keskkonnategurid muutuvad, jätkub sama faas – kas gametofüüt või sporofüüt. Sellist tsüklit nimetatakse heteromorfseks, millel on ebaregulaarne muutus arenguvormides. Seda märgitakse Mastokarpovide seas. Esinevad isomorfsed tsüklid, mille arenguvormid muutuvad korrapäraselt, mil seksuaalseid ja aseksuaalseid vorme esindavad väliselt identsed iseseisvad vabalt elavad põlvkonnad. See arendustsükkel on tüüpiline Gracilaria, Chondrus, Masella.
Taksonoomia
Rhodophyta osakond on traditsiooniliselt jagatud kahte klassi: Bangioceae - Bangiophyceae ja Floridaaceae - Florideophyceae. Viimasesse klassi kuulub enamik sarlaki perekondi ja liike.
Suurel sügavusel, kuni 250 meetrit, kasvavad nad punased vetikad, nimetatakse teisiti helepunane. Kombinatsioonis korallide ja erksate kaladega loovad erivärvilised sarlakid kalad veealuse maailma ainulaadse ilu. Tegemist on valdavalt suurte vetikatega, kuid näiteks mikroskoopilised punavetikad kuuluvad ka Banguiaceae klassi.
Miks võivad punavetikad kasvada nii märkimisväärsel sügavusel? See küsimus esitati bioloogia ühtsel riigieksamil. Punane pigment võimaldab vetikatel kasvada suurel sügavusel fükoerütriin. Tänu sellele neelavad punavetikad fotosünteesi ajal spektri rohelisi, siniseid, sinakasvioletseid kiiri. Just need kiired, erinevalt punastest, on võimelised tungima sügavale veesambasse.
Punavetikatele on iseloomulik eoste mittesuguline paljunemine, samuti seksuaalne paljunemine(oogaamia), mõnikord toimub vegetatiivne paljunemine talluse osade kaupa.
Punavetikate hulka kuuluvad phyllophora, porphyra, gracillaria, ptilote, chondrius ja kokku on neid umbes viis tuhat liiki.
Lilla- kuni poolemeetrise läbimõõduga lame ja õhuke ovaalne plaat. Seda iseloomustab ainult seksuaalne paljunemine. Meeste sugurakkudel puuduvad flagellad (sperma). See on arusaadav, kuna suurel sügavusel veesamba all on lipu abil raske liikuda.
Vetikate tähendus
Koht toiduahelates, mõju loodusele
1. Tohutu vetikate mass loob fütoplanktoni isegi sisse arktilised mered aastas on 20-30 miljonit isendit kuupmeetrit vesi. Need on esmased tooted, mis moodustavad toiduahela aluse.
2. Fütoplankton toimib toiduna zooplanktonile (teisesed tooted), mida söövad suured mereelanikud, näiteks vaalad. Huvitav on see, et Thor Heyerdahl proovis Kon-Tiki parvel ekspeditsiooni ajal planktonisuppi ja leidis, et see on üsna maitsev ja toitev.
3. Põhjavetikad pakuvad peavarju kaladele ja mitmesugustele mereloomadele ning on neile ka toiduks. Kelp näiteks söövad hea meelega merisiilikud.
4. Vetikad küllastavad ookeane ja atmosfääri hapnikuga.
5. Kui aga vetikad (näiteks Chlamydomonas) paljunevad massiliselt vee nn õitsemise ajal, langeb hapnikusisaldus selles ja vesi küllastub toksiinidest. Jõeelanikud surevad hapnikupuudusesse.
Tähendus inimestele
1. Vetikaid on paljudes piirkondades toiduna tarbitud tuhandeid aastaid. Eriti populaarsed on pruun- ja punavetikad, nagu pruunvetikas, undaria, porphyra, hijiki (ja sargassum üldiselt).
2. Punavetikad on joodiallikad, eriti mõned selle rikkad liigid.
3. Punased vetikad on ka agar-agari allikaks, geeli moodustav aine, mida kasutatakse kondiitritööstuses, bakterite kasvatamiseks jne.
4. Puhastamiseks kasutatakse vetikaid heitvesi fluorist, lämmastikust jne, samuti õhust süsihappegaasist (see on õnnestunud Chlamydomonas, Chlorella, Euglena).
5. Vetikad on toidu lisaained: spirulina, pruunvetikas, fucus, ulva, klorella ja teised.